
方法总结
来自 Bastidas-Ponce 等人 (2019) 的单细胞 RNA-seq 数据,题为“Comprehensive single cell mRNA profiling reveals a detailed roadmap for pancreatic endocrinogenesis”,使用 Scanpy/scVelo/CellRank 工作流进行分析。该数据集 (GEO: GSE132188) 包括 3,696 个细胞,来自 E15.5 小鼠胰腺,分为 8 种已注解的细胞类型,具有剪接和未剪接的计数层 (27,998 个基因预过滤)。
预处理:对基因进行过滤(至少 20 次共享计数),保留了 2,000 个高度变异的基因;使用 30 个主成分和 30 个邻居计算动量。RNA 速度:拟合 scVelo 动态模型 (recover_dynamics, mode='dynamical'),产生速度向量和潜在时间估计。CellRank 命运映射:合并了 80% 的 VelocityKernel 和 20% 的 ConnectivityKernel,并构建了 GPCCA 估计器通过 Schur 分解识别了 8 种宏状态(21 个成分);基于生物学注释设置了 3 个终端状态(Alpha, Beta, Epsilon),并通过吸收概率估计计算了命运概率。通过与命运概率的表达 Pearson 相关性识别谱系驱动基因。使用 GAM 在潜在时间建模基因表达趋势,以谱系特异的命运概率加权计量。
主要发现
分化层次结构和 RNA 速度
RNA 速度流图揭示了明确的单向分化层次结构:导管前体 -> Ngn3 低 EP -> Ngn3 高 EP -> 前内分泌 -> 终端内分泌命运。潜在时间单调进展从 0.0(导管)到 1.0(成熟内分泌)
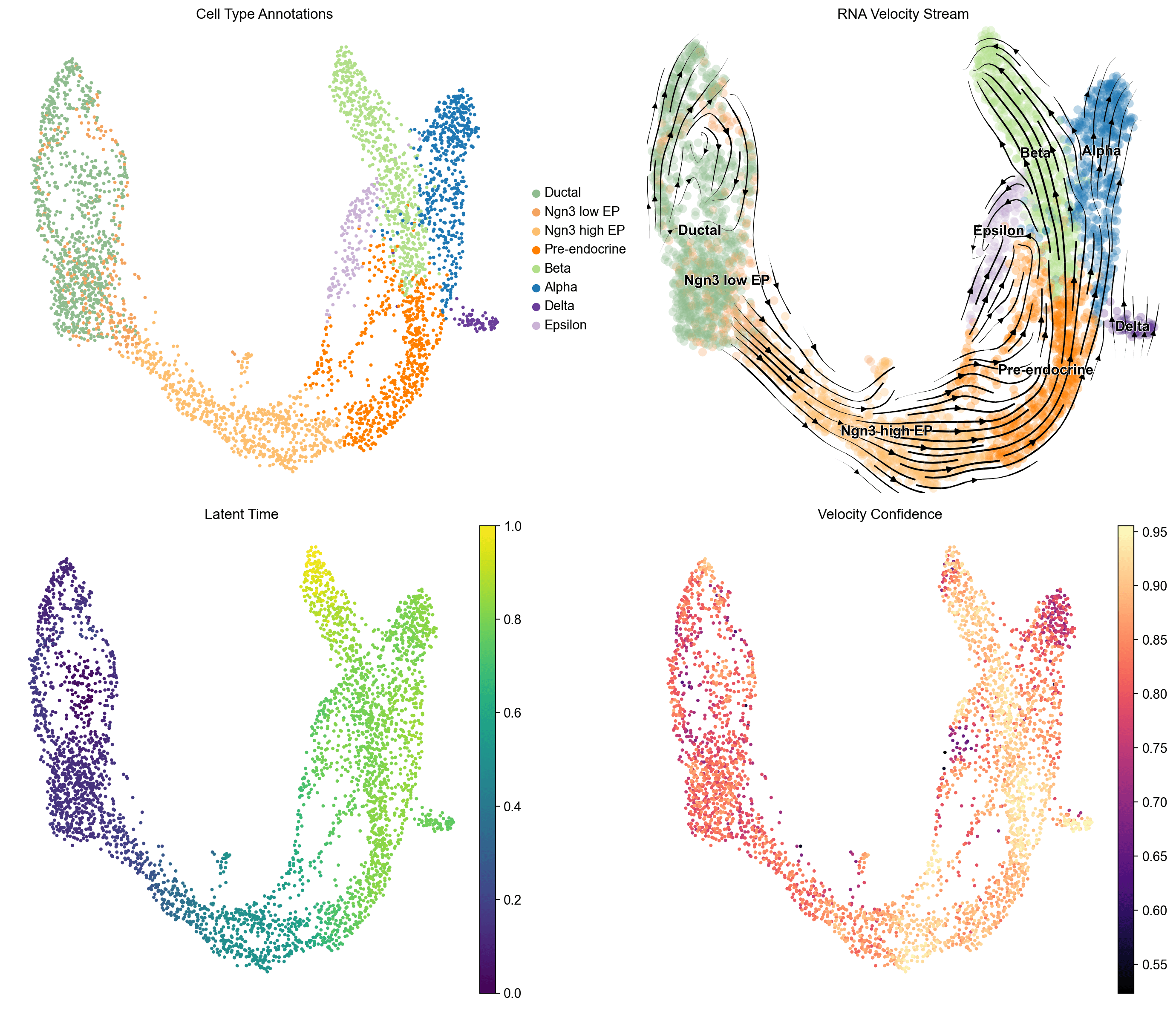
在主要分化轨迹上的速度置信度高(0.80-0.95),验证了方向信号。
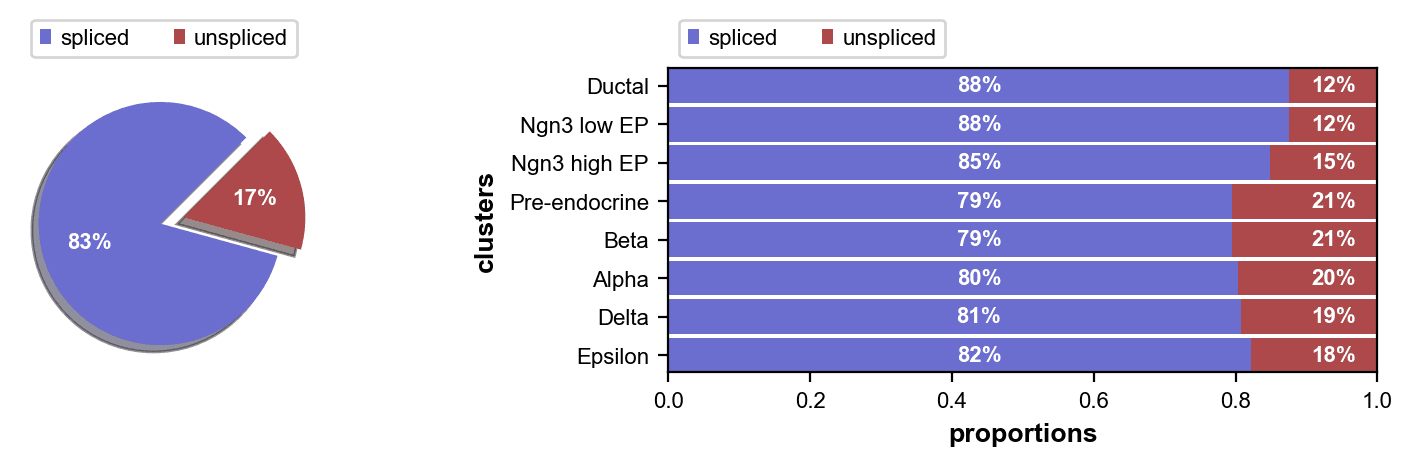
未剪接部分范围从 12%(导管)到 21%(前内分泌,Beta),前内分泌和 Beta 细胞显示出最高的转录更新率
终端状态和命运概率
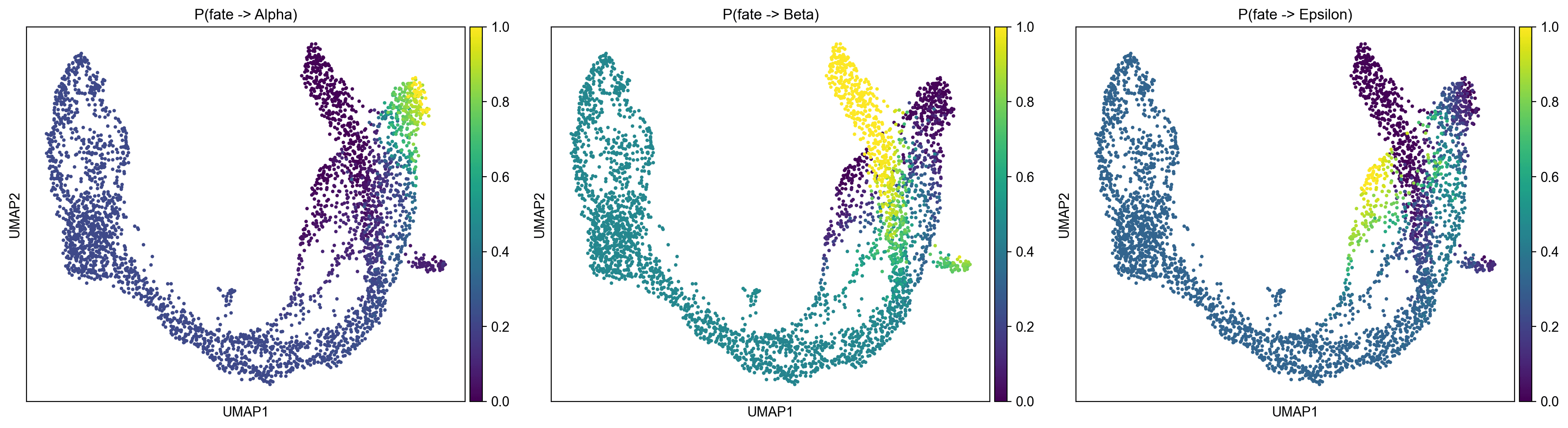
GPCCA 特征值谱在索引 5-6 处显示出间隙,支持 8 种宏状态,干净地映射到已知的细胞群体:4 种导管子状态,Ngn3 低 EP,Alpha(30 个纯 Alpha 细胞),Beta(30 个纯 Beta 细胞),和 Epsilon(30 个纯 Epsilon 细胞)。
Delta 细胞(n=70)由于其小的人口规模和与 Beta 谱系的转录组相近性,未解析为独立的宏状态。
终端状态
平均命运概率(自)
平均命运概率(Delta 向其)
Beta
0.922
0.744
Epsilon
0.893
0.172
Alpha
0.541
0.084
Beta 细胞表现出最强的谱系承诺(P=0.922),表明其具有高度决定论的分化计划。
Delta 细胞主要被分配 Beta 命运(P=0.744),与其已知的发育接近性和共享 Pax4 表达一致。
前内分泌细胞是真正的多能性,三个命运的概率平衡 (P(Beta)=0.477, P(Epsilon)=0.328, P(Alpha)=0.195)。
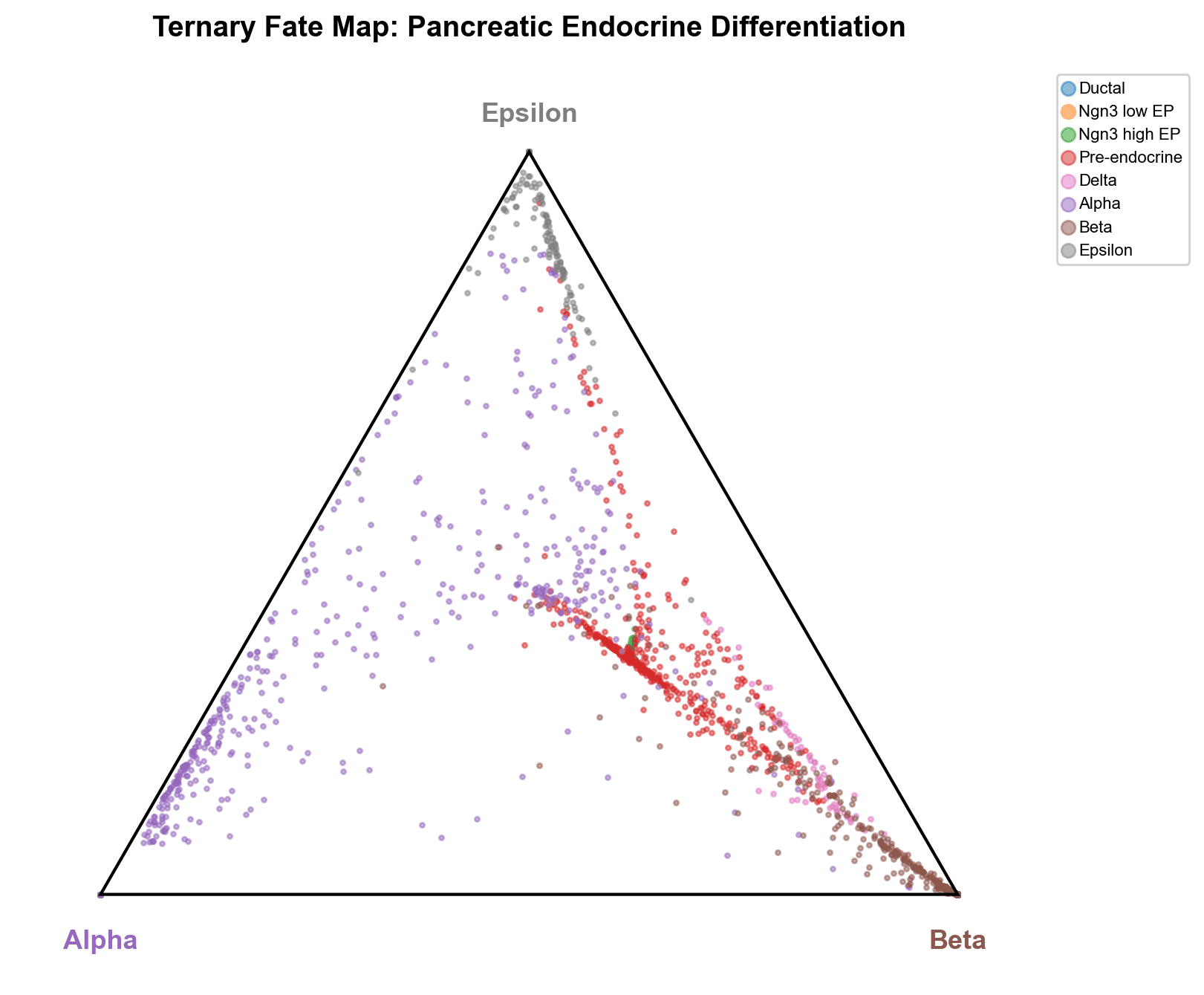
三元命运图投影
简单投影将成熟的 Alpha、Beta 和 Epsilon 细胞放置在各自的顶点位置,而 前内分泌细胞形成一个中间流,向每个终端命运分支。Delta 细胞在沿 Alpha-Beta 边的 Beta 顶点附近形成聚集,强调了它们对 Beta 谱系接近性。
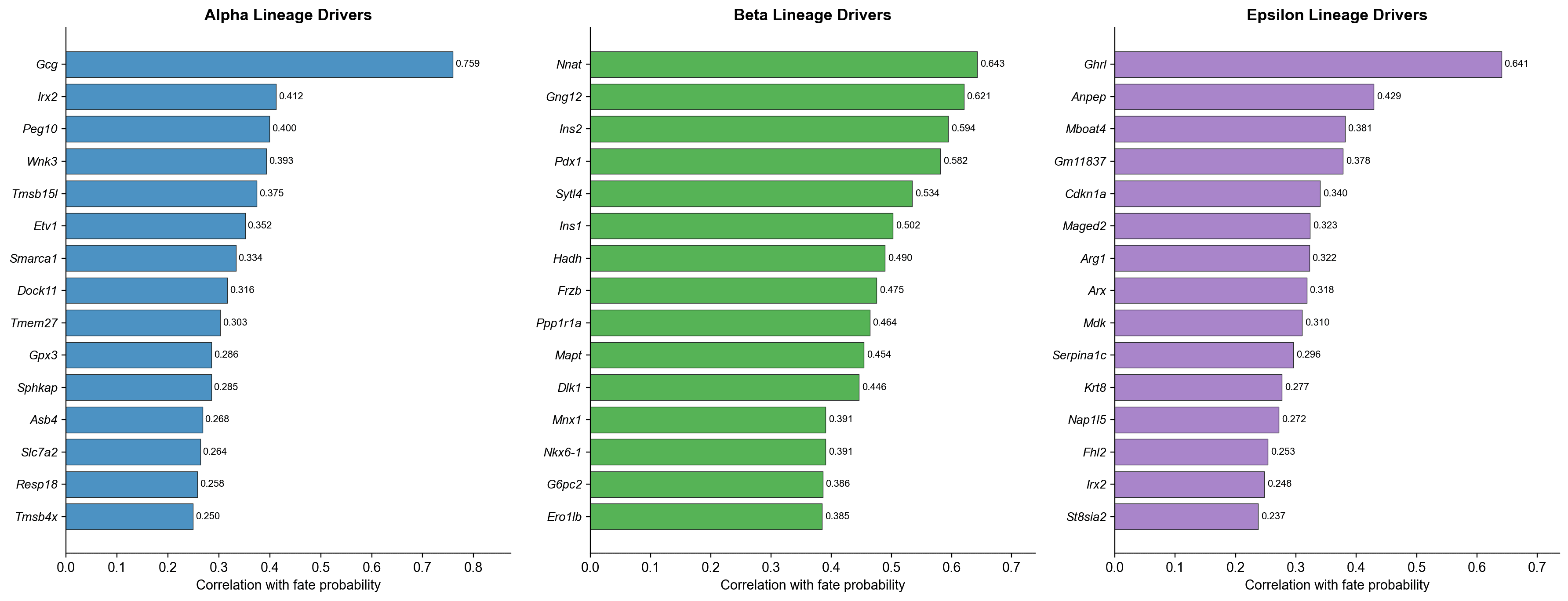
谱系驱动基因
所有排名靠前的谱系驱动者都与已建立的胰腺内分泌生物学匹配良好(所有 q 值几乎为 0):
谱系 | 顶级驱动因素(相关性) | 生物学意义 |
|---|---|---|
Alpha | Gcg (r=0.759), Irx2 (r=0.412), Peg10 (r=0.400), Etv1 (r=0.352) | 胰高血糖素是定义的 Alpha 激素;Irx2 是已知的 Alpha 细胞转录因子 |
Beta | Nnat (r=0.643), Ins2 (r=0.594), Pdx1 (r=0.582), Nkx6-1 (r=0.391) | 胰岛素基因和 Pdx1/Nkx6-1 是典型的 Beta 身份因子 |
Epsilon | Ghrl (r=0.641), Anpep (r=0.429), Mboat4 (r=0.381), Arx (r=0.318) | 饥饿素是 Epsilon 激素;Mboat4(GOAT)是饥饿素酰化所必需的 |
沿拟时间的基因表达动态
Neurog3 在拟时间中期(潜在时间 ~0.4-0.6)显示出特征性瞬时脉冲,标志着内分泌前体规范窗口;这一高峰在所有三个谱系中共享,证实了 Neurog3 作为共同内分泌门户因子的作用
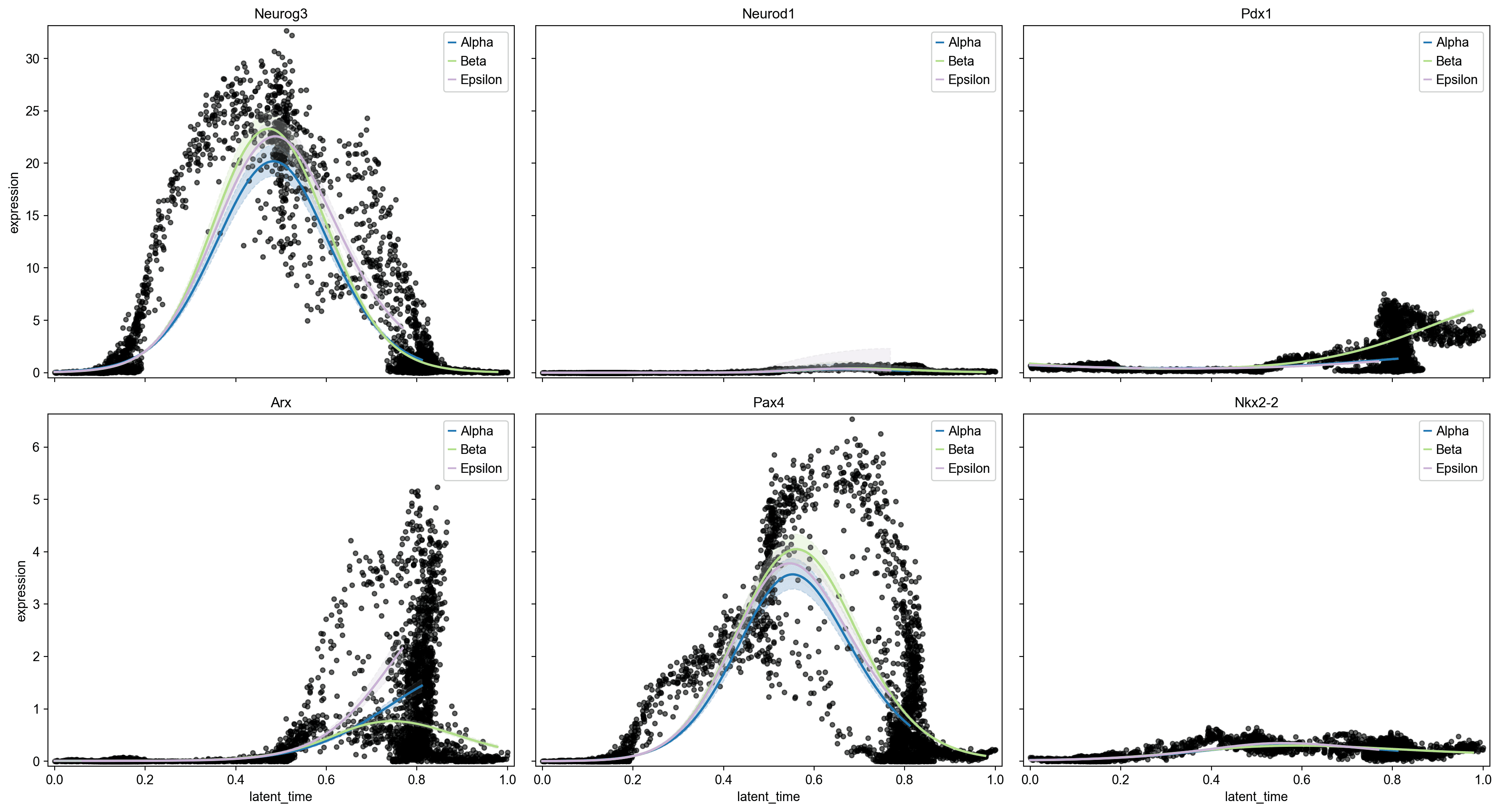
Ins1/Ins2 在 Beta 谱系轨迹上逐渐增加,在晚期拟时间(>0.85)达到最大表达。
Ghrl 在潜在时间 0.65-0.85 窗口中显示出 Epsilon 特异性的上调。
Arx 在 Alpha 谱系上显示出特异性的晚期上调(潜在时间 >0.6),与其作为 Alpha 细胞命运决定因子的角色一致;此时 Alpha 趋势线与 Beta/Epsilon 明显分离。
Pax4 与 Neurog3 同时达到峰值,并在多个谱系中表达,然后解析,与其在 Beta/Delta 命运决定中的已知角色一致。
解释
该分析使用 CellRank 框架提供了 胰腺内分泌细胞命运决定的全面概率重建。一些发现值得强调:
分化是分层的,并且在很大程度上是不可逆的。 RNA 速度向量始终从前体指向终端状态,没有逆向流动的证据。潜在时间梯度平滑单调,支持通过导管、Ngn3 低/高 EP、前内分泌和终端内分泌状态的良好有序的时间进展。
Beta 细胞命运承诺是所有内分泌谱系中最强的。 Beta 细胞的平均命运概率为 0.922,相比之下,Alpha 细胞为 0.541,这表明 Beta 规格涉及一个更加果断的分子开关。这可能反映了涉及 Pdx1、Nkx6-1 和胰岛素基因激活的强正反馈环。
Delta 细胞在转录组上最接近 Beta 谱系。 三元命运投影和吸收概率始终将 Delta 细胞置于 Beta 顶点附近(P=0.744)。这在生物学上与 Pax4 在 Beta/Delta 命运决定中的已知作用一致,并支持一个模型,即 Delta 细胞是从 Beta 承诺谱系内的晚期分支事件中产生的。
Neurog3 脉冲定义了一个重要的规范窗口。 中期 Neurog3 表达的瞬态后期,由 Arx (Alpha) 和 Pax4 (Beta/Delta) 的后续表达发散,重现了内分泌规范的已建立模型,其中 Neurog3 激活下游转录因子,解析为谱系特异性程序。
限制:Delta 细胞(n=70)无法解析为独立的终端状态。Alpha 谱系中 Gcg 的 GAM 趋势未能收敛,可能是因为 Alpha 细胞集中在潜在时间的极端位置,中间采样不足,难以进行平滑曲线拟合。该分析使用了单个时间点 (E15.5);结合原始研究的完整 E12.5-E15.5 时间过程将提供更全面的时间覆盖。
建议的下一步
将 Delta 包含为终端状态,通过增加宏状态分辨率(n_states > 8)或手动定义 Delta 细胞为一个额外的终端状态,以实现独立的 Delta 谱系分析。
应用 CellRank 2 的 RealTimeKernel 结合 GSE132188 数据集的多时间点数据(E12.5-E15.5),使用最佳运输法,捕捉单快照速度分析无法解析的时间动态。
进行差异基因表达在具有高与低命运概率的前内分泌细胞之间的每个谱系(例如,“Beta 向性”与“Alpha 向性”前内分泌细胞)中,识别早期谱系启动特征。
通过 CellRank PseudotimeKernel 交叉验证,作为 VelocityKernel 的替代,评估命运概率估计对方向性信号选择的稳健性。
聚类基因表达趋势(使用
cr.pl.cluster_trends)以识别沿每个谱系共同调控的基因模块,可能揭示驱动内分泌命运决定的新调控程序。