
Structural Biology & Molecular Modeling
Cryo-EM Structural Analysis of Nucleosome Complexes
How Proteins Like OCT4 Open DNA and Histone Modifications Regulate Access
Overview
Understanding how proteins interact with chromatin is critical for target discovery and epigenetic drug development. However, extracting mechanistic insights from structural data across studies remains fragmented and technically intensive.
In this use case, we used Drylab to analyze a panel of nucleosome complexes derived from the Protein Data Bank, including cryo-EM and X-ray structures spanning OCT4 pioneer factor binding, H2B ubiquitination, and chromatin regulatory complexes.
Starting from PDB IDs alone, Drylab performed automated structural alignment, interface quantification, and comparative analysis across complexes — enabling a unified view of nucleosome regulation.
Key Insights
Pioneer factor activity is structurally distinct
OCT4 primarily engages nucleosomal DNA and induces large-scale distortion of the nucleosome, consistent with chromatin opening.Ubiquitination machinery acts locally
The Bre1-Rad6 complex binds specifically to the acidic patch with minimal disruption to nucleosome structure, indicating precise enzymatic targeting.Histone ubiquitination is dynamically positioned
Different ubiquitin conformations variably occlude interaction surfaces, suggesting a regulatory “gatekeeping” mechanism for chromatin access.
Scientific Deep Dive
Structural Dataset and Alignment
A diverse set of nucleosome complexes was analyzed, including:
Canonical nucleosome reference (1KX5)
OCT4-bound nucleosomes (e.g., 7U0G, 8G8E/G)
H2B ubiquitination states (8T3Y/W/T, 8V28)
Chromatin-associated proteins (JADE1 PZP domains, HBO1 MYST complex)
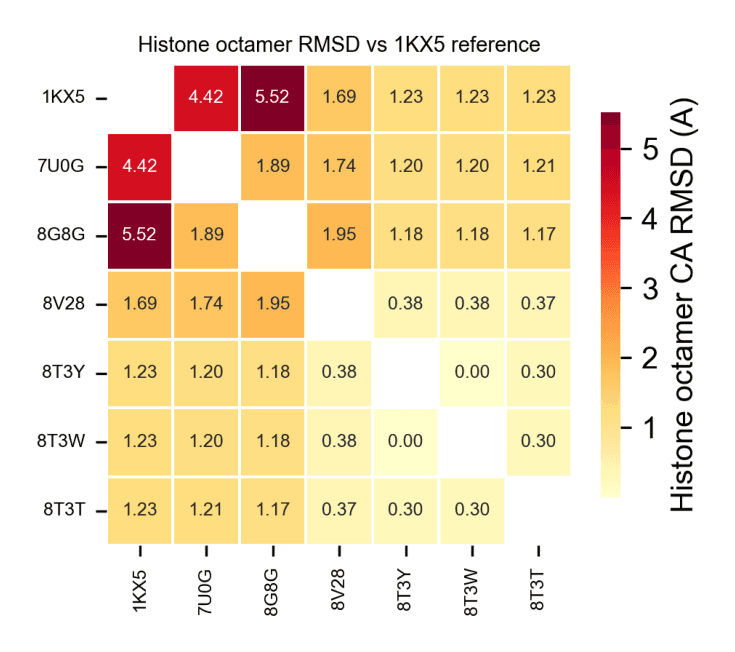
All structures were aligned via Cα superposition to the canonical nucleosome, and structural deviations were quantified using root-mean-square deviation (RMSD).
This analysis revealed two major structural regimes:
A highly conserved cluster (RMSD ~0.001–0.376 Å), comprising ubiquitination-related complexes with minimal perturbation to the histone octamer
A distorted cluster (RMSD ~3.6–4.6 Å), corresponding to OCT4-bound nucleosomes, indicative of significant chromatin remodeling
Protein–nucleosome interactions were systematically quantified using:
Distance-based contact mapping (≤ 5.0 Å cutoff)
Buried surface area (BSA) calculations via solvent-accessible surface area methods
Geometric inference of polar interactions, accounting for the absence of explicit hydrogens in cryo-EM models
OCT4-Mediated Nucleosome Remodeling
OCT4 exhibits a dominant interaction with DNA (BSA ~5,354 Ų) relative to histones (~3,256 Ų), consistent with its role as a pioneer transcription factor. Histone contacts are localized near the acidic patch, where DNA displacement occurs.
OCT4 binding induces substantial conformational changes in the nucleosome:
Elevated RMSD values (up to ~4.65 Å) relative to the canonical structure
Structural features consistent with DNA unwrapping (~25 base pairs) from the histone core
Adoption of a non-canonical POUHD domain conformation, forming a wedge that facilitates chromatin invasion
A notable feature is the positioning of Cys48 (Cys185 in structure numbering):
Highly buried (low solvent accessibility)
Located at the protein–DNA interface (~2.23 Å)
This positioning supports a potential redox-sensitive regulatory mechanism, where oxidation could disrupt DNA binding, consistent with prior functional studies.
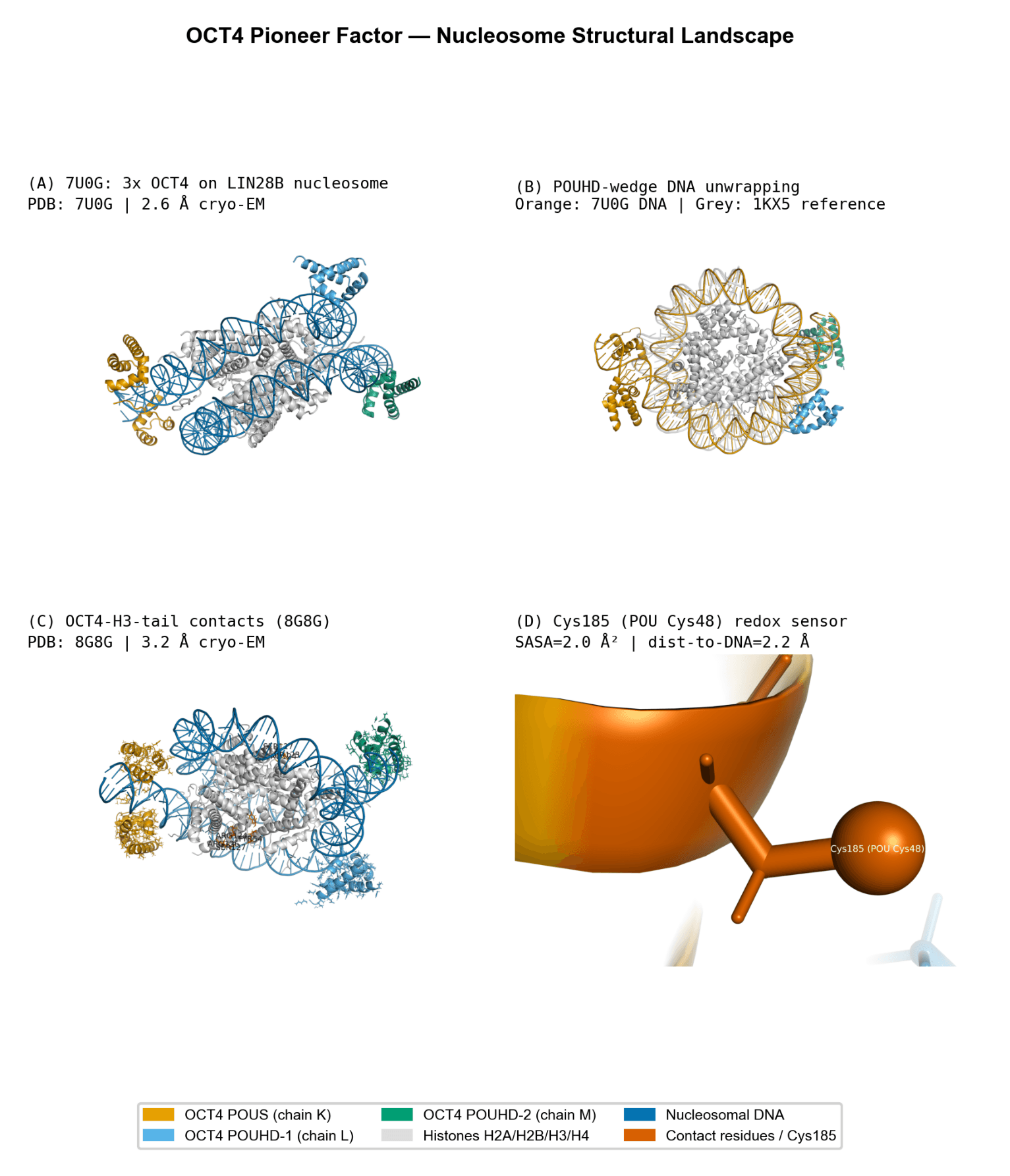
Bre1-Rad6 Ubiquitination Machinery
The Bre1-Rad6 complex engages the nucleosome in a highly targeted manner:
Interaction is concentrated at the acidic patch of histone H2A
Buried surface area is modest (~624–710 Ų), indicating a localized interface
Minimal impact on overall nucleosome conformation
This supports a model in which ubiquitination machinery operates through precision docking, rather than large-scale structural remodeling.
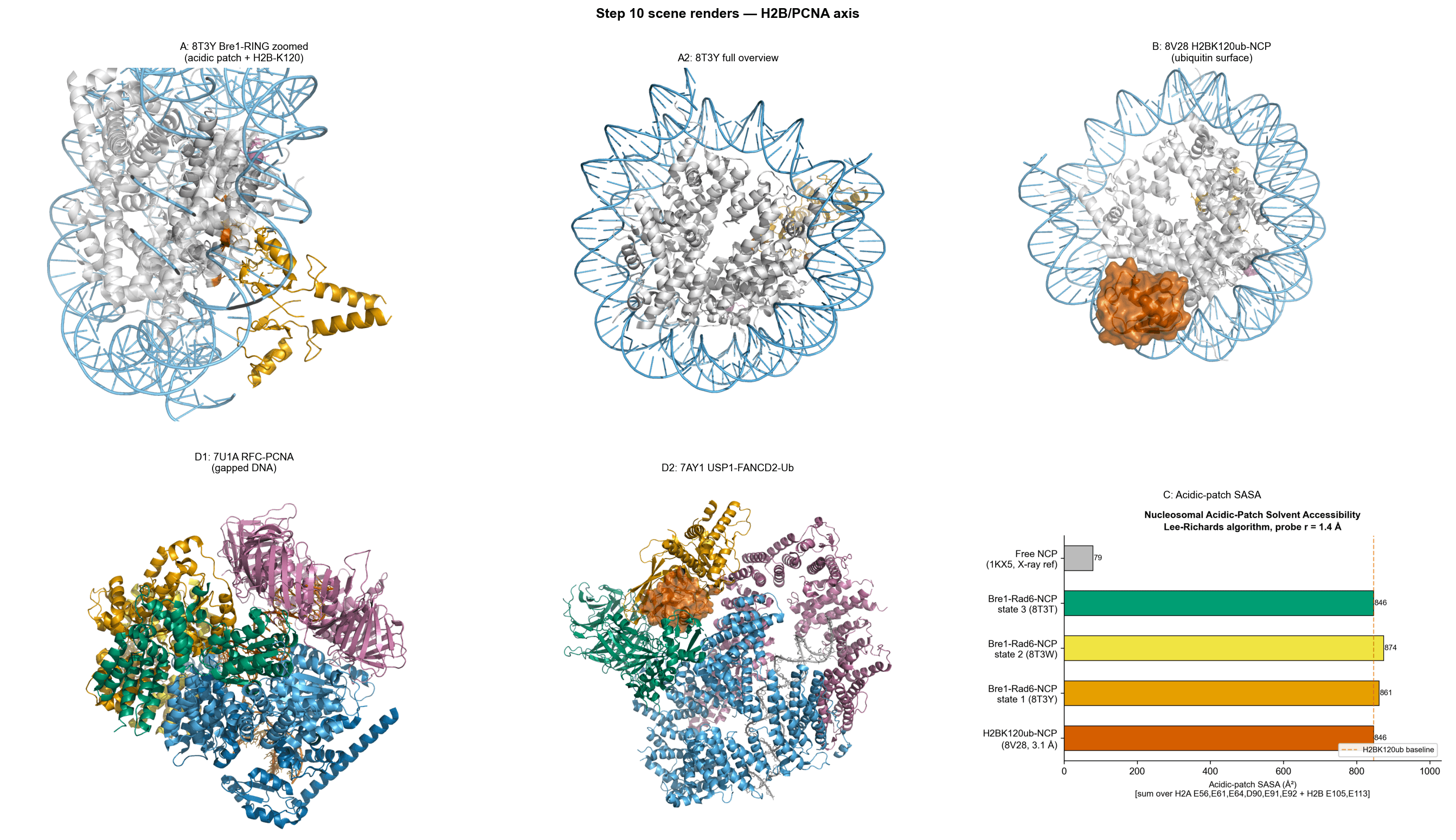
H2BK120 Ubiquitination and Structural Plasticity
Analysis of ubiquitinated nucleosome states reveals:
Significant positional heterogeneity of ubiquitin
In certain conformations (e.g., position 4), minimal interaction with the nucleosome (BSA ~160 Ų)
In other configurations, potential to occlude key interaction surfaces such as the acidic patch
This suggests that H2B ubiquitination functions as a dynamic regulatory element, modulating accessibility for downstream chromatin readers rather than acting as a static structural modification.